М. Лысикова , М. Вальд , З. Масиновски Клиника хирургии 2 го Медицинского факультета Карлова университета, Прага, Чехия; Секции энзимотерапии Общества общей медицины Чешской медицинской ассоциации им. Я.Э. Пуркинье Механизмы воспалительной реакции и воздействие на них с помощью протеолитических энзимов
М. Лысикова , М. Вальд , З. Масиновски Клиника хирургии 2 го Медицинского факультета Карлова университета, Прага, Чехия; Секции энзимотерапии Общества общей медицины Чешской медицинской ассоциации им. Я.Э. Пуркинье Механизмы воспалительной реакции и воздействие на них с помощью протеолитических энзимов
Воспаление относится к древнейшим механизмам защиты. Каскад воспалительной реакции может быть запущен многими стимулами, но в защите организма всегда участвуют пять взаимосвязанных систем: эндотелиальные клетки, тромбоциты, лейкоциты, система коагуляции и система комплемента. Эти системы обладают мощным аутоагрессивным потенциалом, поэтому возможно развитие таких ситуаций, когда воспалительный процесс выходит за рамки защитной реакции, и тогда хроническое воспаление или даже начинающееся аутоиммунное заболевание вызывают альтерацию ткани.
Цель статьи — показать, как системное назначение протеаз животного и растительного происхождения, широко применяемых при лечении воспалительных заболеваний, может эффективно оптимизировать взаимодействие между названными системами. Рассмотрен непосредственный протеолитический эффект, а также роль комплексов протеаз с ? ?2 макроглобулином в связывании цитокинов. (Цитокины и воспаление. 2004. Т. 3, № 3. С. 48–53.)
Воспаление относится к филогенетически старейшим типам защитной реакции организма на различные повреждения и имеет целью устранение причины, локализацию повреждения, удаление поврежденных тканей с последующей регенерацией или восстановлением тканей и восстановлением их функций. Причина, вызывающая воспалительную реакцию, может носить биологический (бактерии, плесени, вирусы, паразиты), физический (излучение, травма), иммунологический (аутоиммунные заболевания), химический или метаболический характер (гипоксия, нарушение метаболизма) [22].
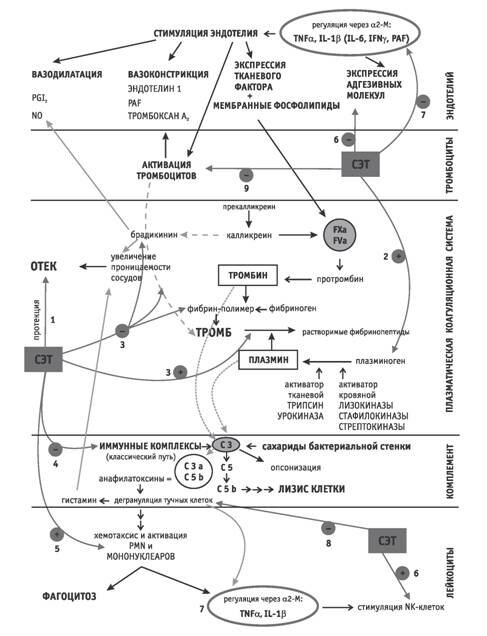
Целью настоящей статьи является обзорный анализ отдельных систем, участвующих в воспалительной реакции организма. Показано, когда и как в регулирование процесса воспаления включаются протеолитические энзимы, содержащиеся в препаратах системной энзимотерапии (СЭТ). Протеазы животного и растительного происхождения при меняют при лечении ряда заболеваний. Кишечнорастворимая форма таблеток рассчитана на доставку энзимов в тон кую кишку, где происходит их всасывание. После всасывания они связываются с белками плазмы, например с ингибитором протеаз ?2 макроглобулином (?2M), что снижает их антигенность, уменьшает скорость распада и выделение с мочой и желчью. В комплексах «протеаза–антипротеаза» энзимы защищены от распознавания иммунной системой и могут быть доставлены в область воспаления. Благодаря этой связи осуществляется также большинство ключевых фармакологических воздействий. Кроме того, экспериментально доказано, что после перорального приема протеаз происходит стимуляция выработки ?2M и изменение его свойств, касающихся, прежде всего, характера его связи с цитокинами [9]. При развитии воспалительной реакции всегда взаимодействуют эндотелий, тромбоциты, лейкоциты, коагуляционная система плазмы и система комплемента. Учитывая аутоагрессивный потенциал некоторых медиаторов, продуцируемых упомянутыми системами, необходимо, чтобы воспалительная реакция—как защитная реакция организма—протекала в темпе и объеме, соответствующем степени повреждения. Важнейшим условием этого является целенаправленное взаимодействие между вышеуказанными системами. Дисрегуляция и делокализация являются решающими факторами развития аутоагрессивного воспаления. Основным средством межклеточных взаимодействий являются цитокины и адгезивные молекулы. Спектр адгезивных молекул на поверхности клетки зависит от того, активирована ли иммунокомпетентная клетка или нет. После активации клетки на ее поверхности начинается экспрессия иных адгезивных молекул, чем в состоянии покоя; благодаря этому активированная клетка способна к новым контактам и может взаимодействовать с другими клеточными системами, что приводит во многих случаях к дальнейшему продуцированию различных типов цитокинов. Протеазы способны непосредственно активировать некоторые клетки, участвующие в воспалительной реакции. В опытах in vitro в присутствии протеаз уже через 10 мин происходило семикратное увеличение активности макрофагов; активность натуральных киллеров (NK клеток), которые способны разрушать опухолевые или инфицированные вирусом клетки без предварительной сенсибилизации, повышалась в 13 раз [17, 18]. Desser с сотрудниками [3] в опытах in vitro и exvivo показала, что у пациентов, получавших peros комбинацию протеаз (препарат Вобэнзим), происходило значительное увеличение продукции мононуклеарами TNF?, IL1?, IL6 — цитокинов, участвующих в острой фазе клеточной реакции. Одновременно в гранулоцитах наблюдалась значительная интенсификация «респираторного взрыва» [28]. Такое «стимулирующее» действие протеаз является важным и необходимым в начальных фазах воспалительной реакции. Процессы активации клеток и усиления продукции цитокинов способствуют защите организма и желательны на начальных фазах воспаления, однако положительная роль этих процессов становится проблематичной в тот момент, когда степень активации перестает быть адекватной и когда первоначально защитный механизм перерастает в патологический процесс (чрезмерная экспрессия некоторых типов адгезивных молекул наблюдается, например, при иммунопатологических заболеваниях). Примером отрицательного влияния перепроизводства цитокинов является гиперпролиферация фибробластов и последующий фиброз тканей вследствие повышенной концентрации TGF?—это типичный пример перехода воспалительного процесса в хроническое течение [23]. Модуляция протеолитическими энзимами активности ?2M приводит к снижению чрезмерно повышенных уровней TGF? у пациентов с фиброзом [22]. Каскад воспалительных реакций начинается с активации эндотелия (рисунок). Сначала происходит вазоконстрикция, в которой кроме других медиаторов участвует тромбоксан А2, вырабатываемый активированными тромбоцитами. После краткой вазоконстрикции происходит вазодилатация, вызванная в основном NO, который продуцируется эндотелиальной NO синтазой в ответ на стимуляцию эндотелия воспалительными цитокинами (TNF? , IL1, IL6, INF?), фактором, активирующим тромбоциты (PAF) и брадикинином (который одновременно повышает сосудистую проницаемость и, таким образом, участвует в развитии отека). Растительная протеаза бромелаин снижает концентрацию брадикинина в жидкости волдырей, вызванных экспериментальным путем на предплечье у добровольцев [2]. Бромелаин также снижает отек, вызванный арахидоновой кислотой, вероятнее всего, влияя на ее метаболизм. Поэтому уже на ранней стадии воспалительной реакции мы наблюдаем противоотечное действие трипсина и бромелаина и отечно протективное действие папаина [5, 19], в то время как химотрипсин способен уменьшить уже более развитый отек. Стимуляция эндотелиальных клеток вызывает, кроме того, экспрессию фосфолипидов на их поверхности. Отрицательно заряженные мембранные фосфолипиды во взаимодействии с фактором Виллебранда позволяют реализовать адгезию тромбоцитов. На поверхности стимулированных эндотелиальных клеток появляются адгезивные молекулы, которые делают возможной адгезию и последующий переход лейкоцитов в воспалительный очаг. Лейкоциты в физиологических условиях «скользят» по эндотелию; такое движение вызвано слабым взаимодействием между адгезивными молекулами на поверхности эндотелиальной клетки (селектины) и лейкоцита. Благодаря активации эндотелиальных клеток и последующей экспрессии адгезивных молекул другого типа—интегринов [LFA 1 (Lymfocyte Function Antigen), Mac 1 (Integrin 1 на поверхности макрофага), VCAM 1 (Vascullar Cell Adhesion Molecule)] — происходит более прочное прилипание лейкоцитов и их последующее проникновение [опосредованное молекулой PECAM 1 (Platelet Endotelial Adhesion Molecule)] в очаг воспаления. Необходимо отметить, что активация эндотелия и экспрессия указанных адгезивных молекул, вызванная эндотоксинами, гистамином, тромбином, анафилатоксинами, простаноидами или PAF, происходит очень быстро (нарастание от секунд до минут) и продолжается обычно в течение нескольких часов. Напротив, такой же процесс, начатый под влиянием воспалительных цитокинов, происходит более медленно, так как речь идет о процессе, зависимом от синтеза белков, однако эффект такой активации продолжается до нескольких дней [10, 22]. Протеазы способны непосредственно регулировать адгезивные молекулы—эксперименты in vitro доказали чувствительность к протеазам известных молекул CD44, CD45, CD4, CD80, CD62L и других [7]. Эта способность протеаз в значительной степени реализуется уже при адгезии лейкоцитов к эндотелию и их последующем проникновении в очаг воспаления.
Воздействие системной энзимотерапии (СЭТ) на механизмы воспалительного процесса
1 — противоопухолевое действие трипсина, химотрипсина и бромелаина и протективное действие папаина [5, 19]; 2 — повышение концентрации плазминогена плазмы [11]; 3 — снижение образования фибрина и повышение удаления уже образовавшихся тромбов [6]; 4 — повышение клиренса иммунных комплексов, утрата способности иммунных комплексов активировать каскад комплемента [13, 14; 21; 25]; 5 — увеличение продукции цитокинов мононуклеарами [3], стимуляция гранулоцитов [29], стимуляция NKклеток [17, 18]; 6 — снижение экспрессии адгезивных молекул [7]; 7 — снижение уровня ?2макроглобулина [13]; 8 — ингибирование дегрануляции мастоцитов [28]; 9 — уменьшение активации тромбоцитов [20]
Кроме того, селективным отщеплением молекул CD4, CD44 и B71 (костимулирующих молекул) на поверхности Т лимфоцитов можно снизить порог активации этих клеток, что и было показано в эксперименте, проведенном на модели аллергического воспаления у животных (аллергического энцефаломиелита), опосредованного Т клетками: назначение Флогэнзима (трипсин, бромелаин, рутин) предотвращало возникновение болезни [26]. Повышение проницаемости эндотелия, вызванное его активацией, делает доступным субэндотелиально расположенный тканевый фактор (TF), который в комплексе с отрицательно заряженными фосфолипидами активирует плазматический фактор VII и через него фактор Х (фактор Стюарта), что ведет к включению коагуляционной системы плазмы PCS (Plasmatic Coagulation System). Если эндотелий активирован воспалительными цитокинами (TNF?, IL1?), то происходит экспрессия тканевого фактора прямо на поверхности эндотелия, что драматически продлевает быструю тромбогенную реакцию эндотелия. Альтернативный путь развития активации PCS (контактная активация) включает переход прекалликреина в калликреин, который отщепляет от кининогенов вазоактивный пептид брадикинин. Ключевым элементом PCS является тромбин, который переводит растворимый фибриноген в нерастворимый фибрин. Фибрин вместе с активированными тромбоцитами образует микротромбы. Тромбы в физиологических условиях в определенной степени лизируются плазмином, активированным в основном тканевым активатором плазминогена tPA (Tissue Plasminogen Activator), выделяющимся из стимулированных эндотелиальных клеток. С точки зрения иммуномодулирующего влияния энзимов уже в этот момент имеет важное значение повышение под влиянием протеаз клиренса воспалительных цитокинов, за счет их связывания с естественными антипротеазами, прежде всего с ?2М. После присоединения протеазы ?2M «активируется», т. е. переходит из так называемой «медленной» формы в «быструю». Обе формы связывают цитокины, но с разной спецификой: нативный свободный ?2M связывает, в основном, IL6, PDGF и NGF; ?2M, активированный протеиназой,—в основном, IL1, TGF? и TGF? [15]. ?2M может также «активироваться» в результате связывания с такими эндо генными протеазами, как плазмин, в результате чего существенно возрастает его специфическое сродство к важнейшему воспалительному цитокину TNF? [27]. Учитывая стимулирующее воздействие экзогенных протеаз на уровень плазмина в крови (см. далее), этот механизм может вносить свой вклад в общую «активацию» ?2M препаратами СЭТ. После связывания со свободным («медленным») ?2M цитокины сохраняют свою активность (и целенаправленно переносятся), в то время как после связывания с активированным («быстрым») ?2M они теряют активность; кроме того, этот комплекс побуждает макрофаги к экспрессии рецепторов для ?2M, что является первым шагом к удалению цитокинов из циркуляции. Снижением концентрации цитокинов протеазы предотвращают чрезмерную и, главное, длительную активацию клеток, потому что снижение количества циркулирующих цитокинов во многих случаях ограничивает экспрессию адгезивных молекул на клетках и таким образом препятствует хронизации воспаления [9, 16]. Действие цитокинов не ограничено ближайшим тканевым окружением—TNF?, IL1 и IL6 включают в воспалительную реакцию весь организм, вызывают так называемую реакцию острой фазы, которая проявляется в выходе большого количества лейкоцитов из костного мозга, синтезе белков острой фазы: С реактивного белка (CRP), фибриногена, ?2М, ?1 антитрипсина и некоторых составляющих комплемента. Клинически отмечается повышение температуры тела и повышенная утомляемость. По общей реакции организма на воспаление можно отчетливо наблюдать противовоспалительное действие протеаз. После приема протеолитических энзимов наблюдалось, на пример, значительное снижение уровня CRP, фибриногена и ?2М [13]. Снижение концентрации ?2 макроглобулина подтверждает его переход из «медленной» формы в «быструю» и повышенное удаление из циркуляции цитокинов быстрой фор мой ?2М. Ускорение удаления воспалительных цитокинов уменьшает вероятность длительной тромбогенной активации эндотелия, что вместе со снижением концентрации фибриногена и брадикинина, с повышением концентрации плазминогена [1, 11] и снижением агрегации эритроцитов и тромбоцитов [12] составляет механизм фибринолитического действия протеаз [4]. Указанное фибринолитическое действие улучшает реологические свойства крови, что, параллельно со снижением отека, способствует восстановлению микроциркуляции. Улучшение реологических свойств крови и лимфы положительно влияет на поступление антибиотиков и химиотерапевтических препаратов в ткани. Следующим шагом в воспалительной реакции организма является активация комплемента. Тромбин и плазмин, так же как полисахариды клеточной стенки бактерий, активируют комплемент так называемым альтернативным путем. Классический путь активации комплемента начинается с иммунных комплексов. Действие комплемента проявляется воспалительным, опсонизирующим и цитотоксическим эффектами. Цитотоксичность направлена в основном против грамотрицательных бактерий. Воспалительное действие определяют анафилатоксины, которые вызывают дегрануляцию тучных клеток. Кроме того, анафилатоксины (компоненты C5b и C3b) обеспечивают активацию и хемотаксис нейтрофильных гранулоцитов и мононуклеарных фагоцитов, которые обеспечивают защиту от микроорганизмов. Но макрофаги являются также и основными продуцентами TNF? и IL1?, длительное чрезмерное влияние которых ведет к необратимому повреждению тканей. Чаще всего речь идет о капиллярах почечных клубочков (иммунокомплексные нефриты), суставах и коже (крапивница) и т. п. Протеолитические энзимы (в основном папаин и бромелаин) подавляют образование иммунных комплексов и повышают их клиренс. Kunze [14] в опытах in vitro показал, что иммунные комплексы, обработанные протеазами, утрачивают способность связывать С1q и активировать каскад комплемента. Это нарушает порочный круг образования иммунных комплексов [13, 21, 25]. В эксперименте [28] было показано также прямое ингибирование дегрануляции тучных клеток под влиянием трипсина, который снижал выделение гистамина (вызванное протамином) на 90 %. Описанное снижение концентрации гистамина способствует также противо отечному действию протеаз. Лейкоциты, которые попадают в очаг воспаления через эндотелий, кроме своих основных задач—фагоцитоза, представления антигена и продукции антител—также продуцируют воспалительные и противовоспалительные цитокины и поэтому играют главную роль во взаимосвязях между разными типами клеток, участвующими в воспалении. Например, тучные клетки, ответственные за наступление реакции повышенной чувствительности немедленного типа, после стимуляции (например, аллергеном) дегранулируют и высвобождают, наряду с другими физиологическими веществами, сильные вазодилататоры—гистамин и серотонин, затем гепарин, метаболиты арахидоновой кислоты и цитокины TNF? и IL4. Th1 лимфоциты также регулируются цитокинами. Задачей вспомогательных Th1 лимфоцитов является взаимодействие с макрофагами, которым после фагоцитоза микроорганизма необходима стимуляция в форме цитокинов, продуцированных именно Th1 (в основном речь идет о INF? и IL2); только после такой активации запускаются бактерицидные механизмы внутри макрофага. И наоборот, активированный макрофаг на своей поверхности экспрессирует адгезивные молекулы (например, ICAM 1), которые во взаимодействии с IL12 ведут к умножению соответствующего клона и дифференциации его в эффекторные Th1 клетки [8, 24]. При иммунной реакции, связанной с B лимфоцитами, которые после стимуляции антигеном превращаются в плазматические клетки и продуцируют антитела, необходим контакт В лимфоцита с адгезивной молекулой CD40 на поверхности Th2 лимфоцитa и стимуляция цитокинами IL4, IL5 и IL6. Для защиты организма принципиально важен выбор правильного иммунного механизма. Инфекции, вызванные внеклеточными микроорганизмами, подавляются антителами (Th2 ответ), а внутриклеточные патогены—макрофагами (Th1 ответ). Этот выбор находится в прямой зависимости от концентрации двух цитокинов—IL12 (продуцируется стимулированными макрофагами и дендритными клетками) и IL4 (продуцируется базофилами и мастоцитами). Анализ влияния протеаз на спектр адгезивных молекул на поверхности Т лимфоцитов и продуцируемые ими цитокины у пациентов с сахарным диабетом I типа показывает, что протеолитические энзимы способны селективно ингибировать Th1 лимфоциты, в результате чего предупреждается развитие воспаления, направленного против собственной ткани. С точки зрения иммуномодуляции очень важна способность трипсина повышать порог активации Т лимфоцитов благодаря влиянию на адгезивные молекулы CD4, CD44 и B71. Повышенная экспрессия CD4, CD44 и B71 и связанное с ней снижение порога активации Т лимфоцитов наблюдается в очаге воспаления. Оно вызвано стимуляцией INF? и направлено на активацию Т лимфоцитов. С учетом того, что активированный Т лимфоцит также продуцирует INF?, происходит усиление всего процесса. Действие трипсина на указанные адгезивные молекулы (одновременно с повышенной элиминацией INF? через связь с комплексом протеаза—?2М) возвращает T лимфоциты в состояние покоя и этим ограничивает гипер активацию воспалительного процесса [26]. Воздействие протеаз на воспалительный процесс является многосторонним, что позволяет в ряде случаев отказываться от широкого спектра иных противовоспалительных средств. Влияние протеолитических энзимов на ключевые механизмы воспалительной реакции включает в себя и поддержку его в физиологической фазе и эффективное сдерживание при превышении уровней защитного механизма. Наглядным примером удержания динамического равновесия воспалительных и противовоспалительных элементов с помощью экзогенных протеаз является динамика ?2М. С одной стороны, протеазы активируют производство ?2М макрофагами, что отражает их «стимулирующую» роль в начальной (физиологической) фазе воспаления. Одновременно происходит снижение уровня этого «медленного» ?2М в результате его связывания с протеазами и перехода в «быструю» форму с последующей элиминацией макрофагами (вместе с воспалительными цитокинами). Этот процесс становится преобладающим—и, бесспорно, терапевтически выгодным—в условиях пролонгированного воспаления. В отличие от классических противовоспалительных лекарств (например, нестероидных препаратов) протеазы оптимизируют физиологический процесс воспаления и препятствуют переходу процесса в аутоагрессию и хроническое течение.
ЛИТЕРАТУРА
1. Веремеенко К.М. та iншi. Вплив полiензимних препаратiв на систему фiбринолiза // Лаб. Дiагностика. — 2002. — № 1. — С. 10–12.
2. Bodi T. The effect of oral bromelains on tissue permeability to antibiotics and pain response to bradykinin: double studies on human subjects // Clin. Med. — 1966. — Vol. 7. — P. 61–65.
3. Desser L., Sakalova A., Zavadova E. et al. Concentrations of soluble tumor ne crosis factor receptors, ?2microglobulin, IL6 and TNF in serum of multiple myeloma patients after chemotherapy and after combined enzyme chemother apy // Int. J. Immunother. — 1997. — Vol. XIII, № 3–4. — P. 121–130.
4. Eim J. LAVH by electrocoagulation, postoperative period and systemic enzyme therapy // World congress of gynaecologic endoscopy. — Rome, Italy, June 18–22, 1997.
5. Gaggi R., Biaggi G.L. Pharmacologic study of Wobenzym. — Bologna: Insti tuto di farmacologia Publ., 1985.
6. Guggenbichler J.P., Einflu? hydrolytischer Enzyme auf Thrombusbildung und Thrombolyse // Medwelt. — 1988. — Bd. 39. — S. 277–280.
7. Hale L.P., Haynes B.F. Bromelain treatment of human T cells removes CD44, CD45RA, E2/MIC2, CD6, CD7, CDS, and Leu 8/LAM1 surface molecules and markedly enhances CD2mediated T cell activation // J. Immunol. — 1992. — Vol. 149, № 12. — P. 3809–3816.
8. Horejsi V., Bartunkova J. Zaklady imunologie. — Triton, 1998.
9. James K. Interactions between cytokines and alphamacroglobulin // Immunol. Today. — 1990. — Vol. 11, № 5. — P. 163–166.
10. Klaschka F. Systemova enzymoterapie — imunologicke mechanismy — Wobe Mugos E v komplexni protinadorove lecbe. Dum Mediciny.
11. Kleine M.W, Pabst H. Die Wirkung einer oralen Enzymtherapie auf experimentell erzeugte Hamatome // Forum prakt. Allg.Arzt. — 1988. — Bd. 2. — S. 2–8.
12. Koshkin V.M., Kirienko A.I. Systemic enzyme therapy in the treatment of acute throm bosis of superficial veins in the lower extremities and postthrombophlebitic dis ease // Int. J. Immunother. — 2001. — Vol. XVII, № 2–3–4. — P. 121–124.
13. Kunze R., Ransberger K., Buschmans E. et al. The adhesion protein CD44 is modulated by proteolytic enzymes // 2nd Int. Congr. on Biological Response Modifiers. — San Diego, Ca., 1993. — Poster 21.
14. Kunze R., Ransberger K., Stauder G., Gebauer F. Proteolytische Enzyme mod ulieren die C1qBindungskapazitat gebundener Immunkomplexe in vitro // J. Inf. Immunol. Dis. — 1996. — Vol. 1 (Suppl.) — P. 17–29.
15. LaMarre J., Wollenberg G.K., Gonias S.L., Hayes M.A. Cytokine binding and clear ance properties of proteinase — activated alpha2 macroglobulins // Lab. In vest. — 1991. — Vol. 65, № 1. — P. 5612–5622.
16. Lauer D., Muller R., Cott Ch. et al. Modulation of growth factor binding prop erties of alpha2 macroglobulin by enzyme therapy // Cancer Chemother. Phar macol. — 2001. — Vol. 47 (Suppl.) — P. 4–9.
17. Leskovar P. AIDS: Neuartige Therapiekonzepte // Dtsch. Zeitschr. Onkol. — 1990. — Vol. 22. — P. 26.
18. Leskovar P., Zanon P.R., Nachbar F., Meschik M. Die negative Rolle von Immunoko mplexe auf die Immunregulation // Rheuma. — 1993. — Bd. 13, № 6. — S. 1–7.
19. Lindner A., Forster O. Berichtuber den Einflu? von Wobenzym auf verschiedene Rattenpfotenodeme. — Wissenschaftliche Monographie, 1985.
20. Mazurov V.I., Stolov S.V., Linetskaja N.E., Onyschenko E.F.: Systemic enzyme therapy in complex treatment of angina pectoris // Int. J. Immunother. — 2001. — Vol. XVII, № 2–3–4. — P. 113–120.
21. Nakazawa M., Emancipator S.N., Lamn M.E. Proteolytic enzyme treatment re duces glomerular immune deposits and proteinuria in passive Heymann ne phritis // J. Exp. Med. — 1986. — Vol. 164. — P. 1973–1987.
22. Necas E. Obecna patologicka fyziologie. — Praha: Karolinum, 2000.
23. Osterreicher J., Skopek J., Navratil L. et al. Enteral administration of protein ase mixture inhibits development of radiation pneumonitis and lung fibrosis in rats // Int. J. Immunother. — 2000. — Vol. XVI, № 3–4. — P. 45–51.
24. Roep B.O., Engel N.K., Halteren A.G.S. et al. Modulation of autoimmunity to betacell antigens by proteases // Diabetologia. — 2002. — Vol. 45. — P. 686–692.
26. Steffen C., Menzel J. InvivoAbbau von Immunkomplexen in der Niere durch oral applizierte Enzyme // Wien. klin. Wochenschr. — 1987. — Bd. 99, № 15. — S. 525–531.
27. Targoni O.S., TaryLehmann M., Lehmann P.V. Prevention of murine EAE by oral hy drolytic enzyme treatment // J. Autoimmun. — 1999. — Vol. 12. — P. 191–198.
28. Wollenberg G.K., LaMarre J., Rosendal S. et al. Binding of tumor necrosis fac tor alpha to activated forms of human plasma alpha 2 macroglobulin // Am. J. Pathol. — 1991. — Vol. 138. — P. 265–272.
29. Wood G.R., Ziska T., Morgenstern E., Stauder G. Sequential effects of an oral en zyme combination with rutosid in different in vitro and in vivo models of inflam mation // Int. J. Immunother. — 1997. — Vol. XIII, № 3–4. — P. 139–145.
30. Zavadova E., Desser L., Mohr T. Stimulation of reactive oxygen species pro duction and cytotoxicity in human neutrophils in vitro and after oral ad ministration of a polyenzyme preparation // Cancer Biotherapy. — 1995. — Vol. 10, № 2. — P. 147–152.
Mechanisms of inflammatory reaction and proteolytic enzymes influence upon them M. Lysikova , M. Wald1 , Z. Masinovsky2 1 Clinic of Surgery, and Faculty of Medicine, Charles University, Prague, Czech Republic; 2 Sections of Enzymotherapy, Society of General Practice, Czech Medical Association of J.E. Purkyne Inflammation belongs to the oldest defence mechanisms. Although a wide range of stimuli may trigger the cascade of an inflammatory reaction, there are always five systems interacting in the body defense: endothelial cells, platelets, leucocytes, coagulation system and complement system. However, theсe systems have also a strong autoaggressive potential which can lead to the situation when the inflammatory process crosses the barrier of an effective protective tool and the tissue becomes altered by a chronic inflammation or even by an autoimmune disease onset. The aim of this article is to demon" strate how the systemically administered proteases of animal and plant origin, widely used in the treatment of inflammatory conditions, can successfully optimize the interaction between the above mentioned systems. Direct proteolysis and the cytokine binding affinity of ?2 macroglobulinprotease complex are considered. (Cytokines and Inflammation. 2004. Vol. 3, № 3. P. 48–53.)