Лекція 7 Хромосомна теорія спадковості. Зчеплення зі статтю
Хромосоми відіграють велику біологічну роль і мають пряме відношення до передачі спадкових властивостей. Роль хромосом у передаванні спадкової інформації була доведена завдяки: а) відкриттю генетичного визначення статі; б) встановленню груп зчеплення ознак, які відповідають кількості хромосом; в) побудові генетичних, а потім і цитологічних карт хромосом. Обґрунтування хромосомної теорії дано у роботах Т. Моргана К. Бріджеса і А. Г. Стертеванта.
У всіх прикладах схрещування, наведених вище, мало місце незалежне комбінування генів, що належать до різних алельних пар. Це можливо лише тому, що названі гени локалізовані у різних парах хромосом. Проте кількість генів значно перевищує кількість хромосом. Отже, у кожній хромосомі локалізовано багато генів, які успадковуються разом. Гени, локалізовані в одній хромосомі, називаються групою зчеплення. Природно, що у кожного виду організмів кількість груп зчеплення дорівнює кількості пар хромосом, тобто у гороху – 7, у кукурудзи – 10, у томатів – 12, але у організмів, які мають гетеросоми, кількість груп зчеплення буде більше на 1, ніж кількість пар хромосом. Так, у дрозофіли 4 пари хромосом, а груп зчеплення буде 5, у людини 22 пари аутосоми та 2 статеві Х і Y, тому 24 групи зчеплення.
Проте виявилось, що гени, які знаходяться в одній хромосомі, зчеплені не абсолютно. Під час мейозу, при кон’югації хромосом гомологічні хромосоми обмінюються ідентичними ділянками. Цей процес називають кросинговером, або перехрещенням. Кросинговер може відбуватися у будь-якій ділянці хромосоми, навіть у кількох місцях однієї хромосоми. Чим більша відстань між окремими локусами, тим частіше між ними відбувається перехрестя і обмін ділянками.
Морган вдало обрав для дослідження муху-дрозо-філу, яка згодом стала класичним об’єктом для генетичних експериментів. Дрозофіл легко утримувати в лабораторії, вони мають значну плодючість, швидку зміну поколінь (через 1,5 – 2 тижні), невелике число хромосом, що спрощує спостереження.
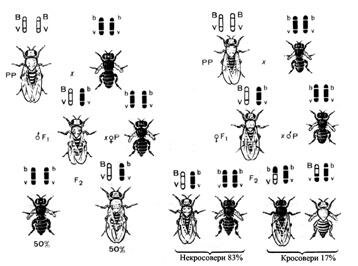
Явище зчепленого успадковування Т. Морган встановив на такому досліді (рис.7).
У дрозофіли гени довжини крил (М – нормальні, м – короткі) і кольору тіла (В – сірий, в – чорний) локалізовані в гомологічних хромосомах, тобто належать до однієї групи зчеплення. Якщо муху, яка має обидва рецесивні алелі (bbmm), схрестити з гомозиготою за домінантними алелями (ВВММ), то в першому поколінні все потомство виявляється гетерозиготним з домінантними виявами ознак у фенотипі (ВbМт). Тут ще немає ніякої відмінності від звичайного дигібридного схрещування. Для виявлення якості гамет першого покоління необхідно провести аналізуюче схрещування, взявши гібридну самку і рецесивного за обома ознаками самця (тобто чорного короткокрилого).
Якщо два гени, які належать до різних алельних пар, локалізовані у різних хромосомах, то дигетерозигота буде утворювати чотири типи гамет: 25% ВМ, 25% Вт, 25% bМ і 25% bm. При аналізуючому схрещуванні такі дигетерозиготи дадуть чотири типи нащадків: сірих довгокрилих, сірих короткокрилих, чорних довгокрилих і чорних короткокрилих у співвідношенні 1:1:1:1.
У нашому експерименті такого співвідношення потомків не буде: В і М знаходяться в одній групі зчеплення, обидва домінантні алелі локалізовані в одній хромосомі, тому гени В і М незалежно один від одного не комбінуються. При абсолютному зчепленні обох генів необхідно очікувати тільки два типи гамет: 50% ВМ і 50% – bm. При аналізуючому схрещуванні має утворитися половина мух сірих довгокрилих, а половина – чорних короткокрилих.
Фактично ж гібридна самка при аналізуючому схрещуванні дає таке потомство:
41,5% сірих довгокрилих;
8,5% чорних довгокрилих;
41, 5% чорних короткокрилих;
8,5% сірих короткокрилих.
Переважання сірих довгокрилих і чорних короткокрилих мух вказує на те, що алелі ВМ і bm тісно зчеплені. Особини таких фенотипів утворюються з гамет, у яких ці хромосоми не перехрещуються. З іншого боку, поява сірих короткокрилих і чорних довгокрилих свідчить про те, що у певній кількості випадків відбувається порушення зчеплення між алелями В і М та b і m. Це наслідок обміну (кросинговеру) ідентичними ділянками хромосом.
Рисунок 7 – Дослід Т. Моргана
На підставі отриманих даних Морган припустив, що гени, які визначають забарвлення тіла і форму крил, розміщені в одній хромосомі, але у процесі мейозу під час утворення гамет гомологічні хромосоми можуть обмінюватися ділянками – відбувається кросинговер.
Обмін ділянками між гомологічними хромосомами має велике значення для еволюції, тому що значно збільшує можливості комбінування мінливості. Внаслідок перехресту добір у процесі еволюції йде не за цілими групами зчеплення, а за групами генів і навіть окремими генами. Адже в одній групі зчеплення можуть знаходитися гени, які кодують поряд з адаптивними (пристосувальними) і не адаптивні стани ознак. У результаті перехресту «корисні» для організму алелі можуть бути відділені від «шкідливих» і, отже, виникнути більш корисні для існування види генів, комбінації генів – адаптивні.
Лінійне розташування генів. Генетичні карти еукаріотів
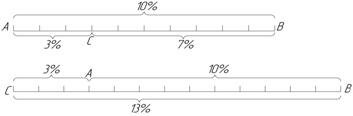
Існування кросинговеру дозволило школі Моргана розробити у 1911-1914 рр. принцип побудови генетичних карт хромосом. У основу цього принципу покладене уявлення про розміщення генів вздовж хромосоми лінійно. За одиницю відстані між двома генами домовились брати 1% перехрещення між ними. Цю величину називають сантиморганідою, на честь генетика Т. Г. Моргана.
Припустимо, що до однієї групи зчеплення належать гени А і В. Між ними виявлене перехрещення у 10%. Отже, ці гени знаходяться на відстані 10 одиниць (сантиморганід). Припустимо також, що до цієї самої групи зчеплення належить ген С. Щоб знайти його місце у хромосомі, треба з’ясувати, який процент перехрещення він дає з кожним з двох уже відомих генів. Наприклад, якщо з А він дає 3% перехрещення, то можна припустити, що ген С знаходиться або між А і В, або у протилежному кінці, тобто А розташований між С і В. Якщо між В і С виявиться перехрещення у 7%, то на хромосомі їх необхідно розташувати у такому порядку, як показано на верхній схемі .
Якщо ж між В і С перехрещення 13%, то розміщення генів у хромосомі має бути таким, як на нижній схемі.
У загальному вигляді цю закономірність можна виразити такою формулою: якщо гени А, В, С належать до однієї групи зчеплення і відстань між генами А і В дорівнює k одиницям, а відстань між В і С дорівнює m одиницям між А і С може бути або k + m або k - m.
Найбільш детальні карти хромосом складені для дрозофіли, яка давно стала класичним генетичним об’єк-том. Із рослинних об’єктів порівняно добре у цьому відношенні вивчені кукурудза і томати, з тварин – кури і миші. Складені карти хромосом людини для всіх 24 груп зчеплення.
Генетичні карти хромосом будують на базі гібридологічного аналізу. Проте, для дрозофіли використовують значно інший спосіб побудови генетичних карт хромосом. Справа в тому, що в клітинах слинних залозах личинок мух знаходяться гігантські (політенні) хромосоми, які перевищують розмір хромосом з інших клітин у 100-200 разів і містять у 8000 разів більше хромонем. Виявилося, що в тих випадках, коли гібридологічним методом виявлялися які-небудь порушення спадкування, відповідні їм зміни мали місце і у гігантських хромосомах. У результаті співставлення генетичних і цитологічних даних стало можливим побудувати цитологічні карти хромосом. Це відкриття підтверджує вірність тих принципів, що були покладені в основу побудови генетичних карт хромосом.